Es gibt einige Ursachen für Schäden im Gewebe
früher Landpflanzen. Das Gewebe zwischen Epidermis und Leitbündel,
Cortex genannt, konnte schon in der lebenden Pflanze teilweise
zerfallen sein, ähnlich wie bei hohlen Bäumen. Unbekannte
Pflanzenfresser nagten Löcher in Sprosse und Sporangien,
um den Inhalt zu fressen.
Löcher
in lebendem Pflanzengewebe können nicht nur durch teilweisen Zerfall
oder Fraß entstehen, sondern auch durch Verdrängen des Gewebes. Als
seltener Zufall sind solche unterschiedlich entstandenen Hohlräume im
gleichen Querschnitt zu sehen: Rhynie
Chert News 117,
Abb.6.
Manche frühen Landpflanzen hatten schon Mittel erfunden, um
Pflanzenfresser abzuwehren: Borsten
an der Oberfläche, Röhren mit
abschreckender Flüssigkeit (?) entlang der Epidermis, oder einen Schutzwall
(?) im Cortex-Gewebe.
Die
Reaktion der Pflanze auf Fraß kann mehr oder weniger intensiv sein. Bei
der frühen Landpflanze in Abb.1 war die Reaktion deutlich,
denn längs der Wand des gefressenen Loches waren
große Zellen entstanden.
Abb.1: Querschnitt von Aglaophyton
mit
einem großen gefressenen Loch, umgeben
von unnatürlich großen Zellen;
wenige schwach sichtbare Querschnitte ummantelter Pilzhyphen, die
später in der wassergefüllten Höhle gewachsen waren;
waagerechte Grenzflächen wässriger Suspensionen; achat-artige
Wandbeläge und Quarzkristalle, gewachsen
im verbliebenen Hohlraum. Bildbreite 6mm.
Abb.2 (unten): Querschnitte von Aglaophyton
als
hohle Halme: oberer Halm mit Loch in der Wand
und anschließendem Tunnel mit fäulnisresistenter Wand, übriger
Raum mit achat-artigen Auskleidungen; unterer Halm flach gedrückt,
anscheinend
mit ähnlichem Gebilde wie oben, aber kleiner, zufällig den oberen Halm
berührend. Bildbreite 7mm.
Im zunächst unübersichtlichen Gebilde in
Abb.2 ist eine auffällige
Reaktion der lebenden Pflanze auf den Angriff eines Pflanzenfressers
erkennbar. Die nahe Umgebung des (im Bild von unten) in den Spross
gefressenen Tunnels wurde offenbar mit Fäulnisresistenz ausgestattet,
um den Schaden zu begrenzen, der sich andernfalls durch das Gewebe
schnell
ausgebreitet hätte. Das ist ähnlich wie in
Abb.1, wo die neu gewachsenen großen Zellen anscheinend verhindert
haben, dass Fäulnis vom Rande des gefressenen
Loches ausgehend sich ausbreiten konnte.
Als das Cortex-Gewebe in Abb.2 später zerfiel, blieb die resistente
Umgebung des Tunnels als unförmiger breiter Schlauch mit dicker Wand übrig und
verkieselte.
Neu gewachsene Zellen
zwecks Reparatur eines gefressenen Loches sind (bei wenig höherer
Vergrößerung) in Abb.3 zu sehen.
Abb3 (rechts): Zellwachstum zwecks Füllen eines Loches.
Bildbreite 2mm.
Es ist noch unverstanden, warum Aglaophyton
manchmal, aber nicht immer, eine fäulnisresistente Schicht,
2-3 Zellen dick, unter der oft unauffälligen oder zersetzten
Epidermis erkennen lässt (Abb.2). Das meist
dunkle oder schwarze Aussehen der Wände der resistenten Zellen
ist anscheinend ein Mikrobenbelag, der gelegentlich abblättern kann.
Abb.4
(rechts unten): Loch in der fäulnisresistenten Außenschicht, repariert
mit einer 2mm breiten fäulnisresistenten Kuppel.
Von einem besonderen Fall von Schadensbegrenzung wurde in Rhynie
Chert News 60,
Abb.4, berichtet, hier als Abb.4 übernommen:
Die Kuppel muss in der lebenden Pflanze gebildet worden sein, als
das jetzt verschwundene Cortex-Gewebe noch vorhanden war, denn sie besteht aus fäulnisresistent gemachten
Cortex-Zellen. Das ermöglicht Schlussfolgerungen bezüglich des
Phänomens der "hohlen Halme". Diese wurden in [1] als bloßer
Diffusionseffekt gedeutet, wo Kieselsäure nur bis in geringe Tiefe
eindiffundiert und nur dort das Gewebe konserviert. Diese Deutung wurde
widerlegt.
Eine
andere Deutung folgt aus Abb.2,4: In der lebenden Pflanze wurde ein
dünne Cortex-Schicht unter der Epídermis auf unbekannte Weise
verändert, wahrscheinlich als Schutz
für das Gewebe darunter. Diese Randschicht war fäulnisresistent
und deshalb später, als das meiste Gewebe verschwunden war, gut
sichtbar.
Aus unbekanntem Grunde bekam die Randschicht in Abb.4 ein Loch, was für
die
Pflanze anscheinend so gefährlich war, dass sie das Loch mittels einer
Kuppel aus modifiziertem Cortex-Gewebe
überbrückte. Zusammenfassend ist festzustellen, dass die Pflanze
fähig war, auf drohende Gefahren und Schäden flexibel zu reagieren,
hier durch Modifizierung von Cortex-Gewebe
zu speziellen Schichten (Abb.2,4) oder durch verstärktes
Zellwachstum (Abb.1,3).
Der Begriff "hollow straw" ("hohler Halm"), der in der Literatur für Aglaophyton mit
fäulnisresistentem Rohr
benutzt wird, suggeriert den Gedanken bewahrter mechanischer Festigkeit
trotz des Zerfalls des übrigen Cortex-Gewebes. Ein
kleines Loch im Halm vermindert dessen Festigkeit kaum,
weshalb die sorgfältige Bedeckung des Loches in Abb.4 nicht
wegen der Festigkeit erfolgt sein konnte. Deshalb bleibt nur die
Deutung, dass die resistenten Rohre, oft unter der Epidermis von Aglaophyton und
immer im Inneren von Ventarura,
weniger der Festigkeit als dem Schutz
vor Schädlingen (Mikroben, Tiere ?) dienten.
Es kann hier nur vermutet werden, dass
die resistenten Rohre in
Aglaophyton und
Ventarura
die Schädigung des übrigen Gewebes verzögerten
oder verhinderten. Es bleibt ungeklärt, ob der Zerfall des Gewebes in
Abb.2-4 in der lebenden oder in der abgestorbenen Pflanze
erfolgte.
Fundstücke: Abb.1: Rh6/9.2 (2002), Abb.2: Rh15/82.4 (2014 obtained
from Barron),
Abb.3: Rh2/68.1 (2002 obtained from J. Shanks),
Abb.4: Rh12/162.2 (2007).
H.-J.
Weiss 2017
[1] C.L. Powell, N.H. Trewin,
D. Edwards: Palaeoecology and plant succession in a
borehole through the Rhynie cherts, ...
Geological Society, London,
Special Publications 180 (2000), 439-457.

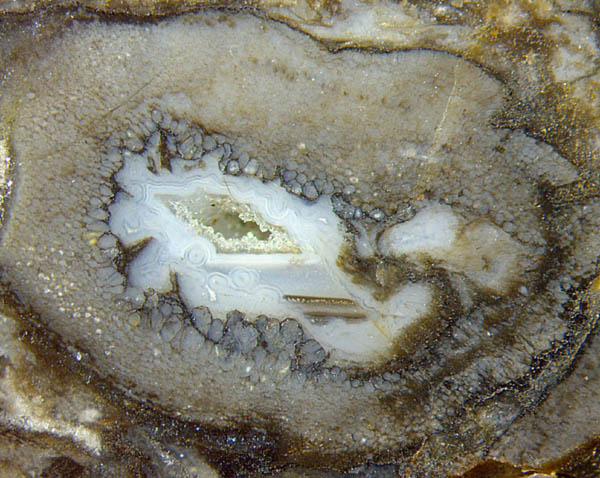
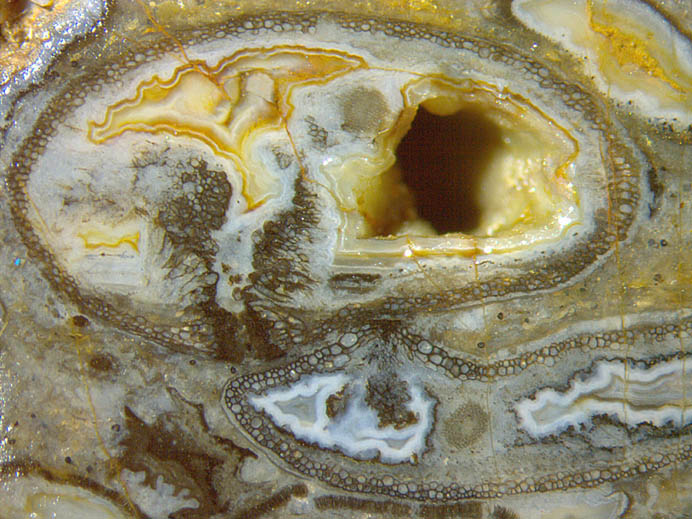
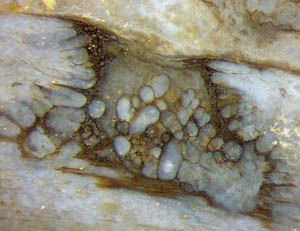
 Abb.4
(rechts unten): Loch in der fäulnisresistenten Außenschicht, repariert
mit einer 2mm breiten fäulnisresistenten Kuppel.
Abb.4
(rechts unten): Loch in der fäulnisresistenten Außenschicht, repariert
mit einer 2mm breiten fäulnisresistenten Kuppel.