

| English version |
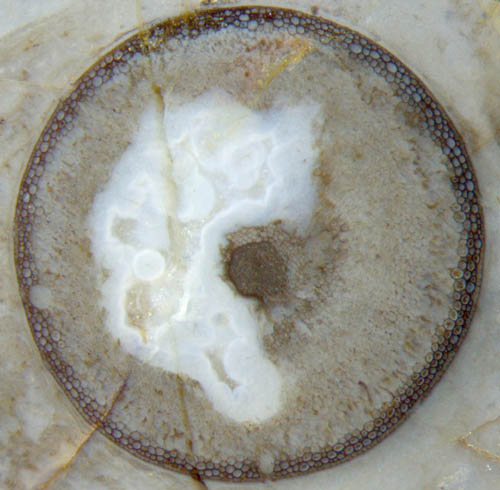
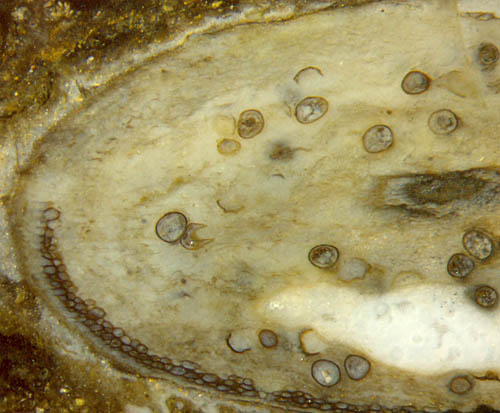
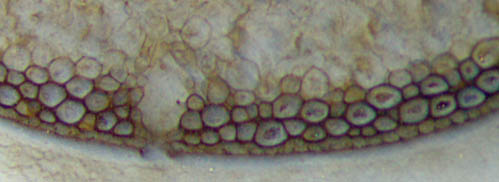 Abb.2:
Teil der Wand von Aglaophyton in
Abb.1, Epidermis links schlecht erhalten. Bildbreite 1.4mm.
Abb.2:
Teil der Wand von Aglaophyton in
Abb.1, Epidermis links schlecht erhalten. Bildbreite 1.4mm.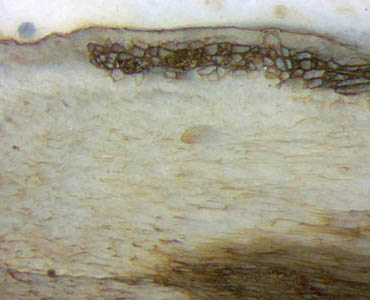
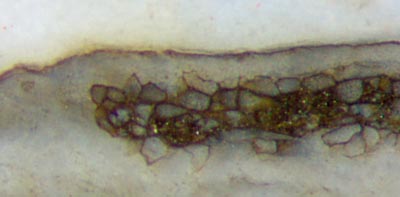
 In
diesem Zusammenhang ist es interessant, dass Trichopherophyton,
obwohl verwandt mit Ventarura, keinen
solchen Ring hat, außer in seltenen Fällen, und dann ist der Ring wie
bei Aglaophyton an der
Epidermis anliegend (Abb.6).
In
diesem Zusammenhang ist es interessant, dass Trichopherophyton,
obwohl verwandt mit Ventarura, keinen
solchen Ring hat, außer in seltenen Fällen, und dann ist der Ring wie
bei Aglaophyton an der
Epidermis anliegend (Abb.6). |
|
97 |
|
|
|
|