Selective decay resistance of soft
tissue in early land plants (3)
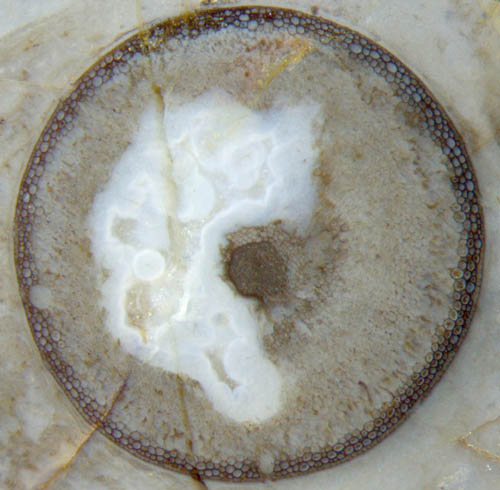
Cross-sections of early
land plants in the Lower Devonian Rhynie chert,
most often Aglaophyton,
may appear as
conspicuous peripheral rings with well-preserved cellular structure
while most of the tissue is severely degraded or no more there at all
(Fig.1). It is
emphasized that the phenomenon is more complex than previously
assumed [1,2]. It
is
described and analyzed here, as done before in Rhynie
Chert News 60,
66,
with the intention that this, combined with more fossil evidence to be
discovered, may eventually lead to an explanation.
Fig.1: Aglaophyton
cross-section 4mm, "hollow straw" aspect with degraded cortex
tissue.
The
"hollow straw" is not really hollow here but all the more informative.
Here again the conspicuously different aspects of the ring and the
larger part of the cortex gives rise to wonder. It has been stated in
the previous contributions that the thickness of the well-preserved
ring is not determined by some diffusion depth of silica from outside
into the plant lying in the silica-rich water. One argument is based on
the observation that most often the epidermis is poorly preserved or
missing (Fig.2). Another argument is provided by
small patches of phloem
seen adjacent to the xylem in Fig.1. The idea of silica
diffusion into
a small depth while all other soft tissue decays [1,2] is
not compatible with the preserved
phloem tissue.
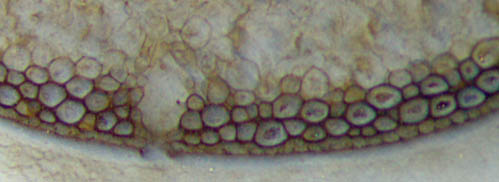
Fig.2: Detail of Aglaophyton "hollow
straw" in Fig.1, epidermis seen poorly preserved below left, better
preserved on the right.
Width 1.4mm
Figs.3,4 (below): Aglaophyton
inclined section: xylem (below), phloem, cortex, epidermis largely
decayed, and persistent
fraction of cortex tissue non-typically arranged as a chute-like strip.
Width of Figs. 2mm, 1mm.
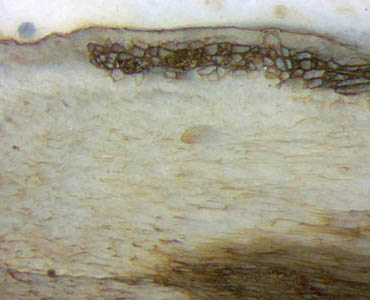
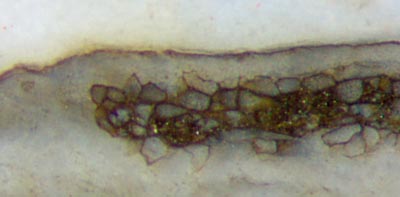
Obviously that idea is also not compatible with
configurations like the one in Figs.3,4, where the small fraction of
persistent tissue,
usually forming a tube and seen as a ring on sections, forms a kind of
chute, seen on sections as a U-shaped strip. The
unexpectedly
distinct border of the strip seen here seems to defy an
explanation.
Chute-like strips of persistent tissue are rare exceptions in
Aglaophyton
but are often seen in Ventarura.
It has to be found out whether the similar aspect is merely incidental
or hints at a deeper connection.
Fig.5 (below): Aglaophyton
inclined section: epidermis and cortex
decayed except for a chute-like persistent
cortex strip. Width of the picture 5mm.
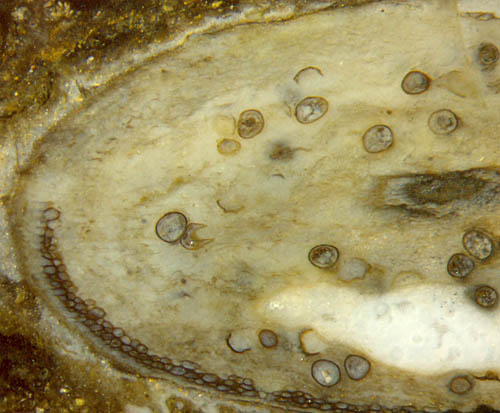
Another one of the irregular Aglaophyton "hollow
straws" formed as a chute-like strip
(Fig.5) shows the peculiar sudden transition from cells with apparently
strong and solid walls to no cells at all, similar to but not quite as
impressive as in Figs. 3,4. The numerous fungus
chlamydospores
indicate that the cortex tissue had been consumed by rot due to some
fungus. The latter can be assumed for the specimen in Fig.3, too, where
the abundant fungus hyphae grown along the cortex are faintly visible
as
streaks. The presence of fungi is also seen in Fig.1 where hypha
cross-sections appear as tiny dark dots, and fungus-infested cortex
cells are seen with brown fill.
As mentioned above, rings of persistent tissue, including chute-like
ones, are typical features of Ventarura,
where they appear on sections of the upper parts of the plant. They are
placed nearly always deep inside the cortex, well away from
the decayed epidermis. Hence, they differ distinctly from the
persistent rings of Aglaophyton,
which are always immediately below the (decayed)
epidermis.
 In
this connection it is interesting that Trichopherophyton,
a relative of Ventarura, usually
does not show such ring, but when it does on rare occasions, it is of
the Aglaophyton
type, adjacent to the epidermis (Fig.6).
In
this connection it is interesting that Trichopherophyton,
a relative of Ventarura, usually
does not show such ring, but when it does on rare occasions, it is of
the Aglaophyton
type, adjacent to the epidermis (Fig.6).
Fig.6 (left): Trichopherophyton,
slightly inclined section with the typical pointed bristles, seen here
on the left. Width of the picture 2.8mm.
As a peculiarity of this Trichopherophyton
section, the cortex appears with two starkly differing aspects: a patch
of "normal" tissue, faintly seen
on the right of the xylem, and conspicuous cells with
apparently strong walls arranged
in a chain-like way along
the epidermis on the left.
Several
observational facts indicate that the fraction of the cortex
tissue usually seen as a ring on cross-sections became
persistent by some modification of the living cells. This
is convincingly demonstrated by the repair of
the ring after damage, which must have been done by the live plant: see Rhynie
Chert News 60,
Fig.4. This indicates that the small fraction of persistent tissue must
have
been of some importance in the life of the plant.
One may wonder how the plant managed to
distribute
the persistence among the cortex cells in a highly selective and well
defined way. The idea suggests itself that cortex cells close to the
epidermis in Aglaophyton are
somehow
predisposed to forming a persistent tube
while
the other cortex cells are prone to decay. This may well be the case
but there must be something else involved, as it is obvious from
Figs.3-6, where the rows of persistent cells end more or less suddenly.
Particularly puzzling is the end of the strip in Figs.3,4. It looks as
if the distribution of rot resistance had not been governed by radial
symmetry but by some process spreading from cell to cell along the
periphery. This applies also to Ventarura,
where the predisposition of
a fraction of cortex does not concern a peripheral tube (or ring in
cross-sections) but a concentric tube well away from the periphery.
In the above pictures, the persistent tissue is dark and therefore
distinctly seen. It must be mentioned that the dark aspect of
the persistent tissue is
not an intrinsic property but a secondary effect mostly but not always
there, which is not considered here as it had been discussed briefly in
Rhynie
Chert News 60.
Finally it can be stated that the persistent fraction of cortex tissue,
normally ring-shaped on cross-sections, often
but not always stained dark,
- is not a result of silica diffusion in Aglaophyton as
assumed in [1,2],
- is no sclerenchyma in Ventarura as
assumed in [3],
- had been predetermined in the live plant by
making part of the cortex tissue rot-resistant,
- poses the problem of how the rot resistance had
been distributed in the cortex in a highly selective way.
H.-J.
Weiss
2016
[1] C.L.
Powell, N.H. Trewin, D. Edwards: Palaeoecology and plant
succession in a borehole
through the Rhynie cherts, ...
Geological Society, London,
Special Publications 180 (2000), 439-457.
[2] www.abdn.ac.uk/rhynie, Chapter Taphonomy.
[3] C.L.
Powell, D.
Edwards,
N.H. Trewin: A new vascular plant from the
Lower Devonian Windyfield chert, Rhynie, NE Scotland.
Trans. Roy. Soc. Edinburgh, Earth Sci.
90(2000 for 1999), 331-349.
 |
 |
97 |